商机详情 -
美国克隆纺锤体卵质量评估
在有丝分裂中,纺锤体的形成与功能至关重要。首先,在有丝分裂前期,中心体复制并分离至细胞两极,形成纺锤体的两极。随后,微管从两极向中心区域延伸,形成纺锤体的主干。在中期,染色体在纺锤丝的牵引下,自动在赤道板排列整齐。当细胞进入分裂后期,纺锤体微管收缩,将染色体牵引至两极,形成两组数目相等的姐妹染色单体。这一过程确保了遗传信息的准确传递,避免了染色体分离错误导致的遗传异常。此外,纺锤体还决定了胞质分裂的分裂面。在染色体分裂的同时,纺锤体中的一部分微管不随染色体分裂到两极,而是停弛在纺锤体中心,形成纺锤中心体。纺锤中心体的中心区域为两组极性相反的微管交叠区,称为纺锤中心区,它决定了接下来的胞质分裂面。胞质分裂开始于分裂后期的较晚期,一般结束于分裂末期后1-2小时,此期间两个子细胞由中心颗粒体连接。纺锤体通过精确控制胞质分裂面的位置,确保了细胞分裂的对称性和稳定性。纺锤体在细胞分裂后期通过微管切割机制实现染色体分离。美国克隆纺锤体卵质量评估
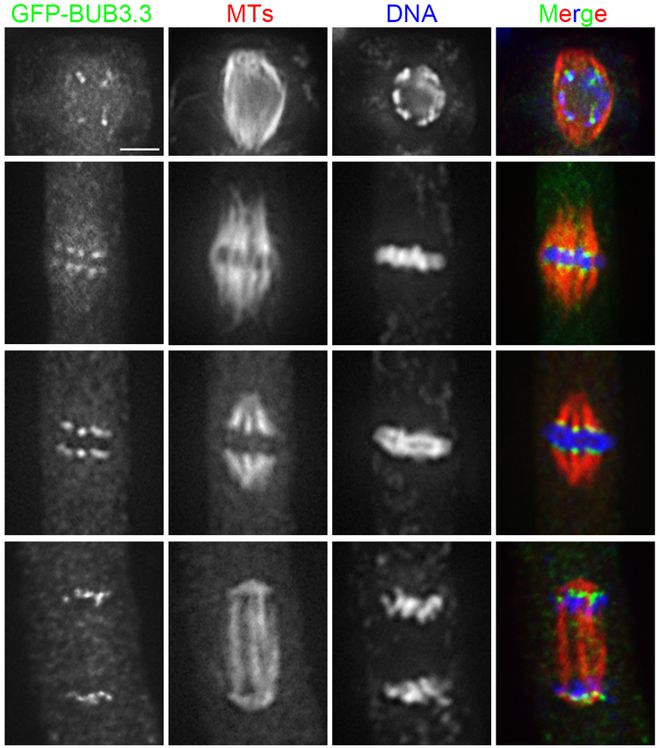
尽管成熟卵母细胞纺锤体冷冻保存技术取得了进展,但仍面临一些挑战。首先,冷冻损伤仍然是制约其临床应用的主要问题之一。尽管玻璃化冷冻法能够在一定程度上减少冷冻损伤,但仍无法完全避免。其次,冷冻保存后的卵母细胞在体外受精和胚胎发育过程中的表现仍存在不确定性。这可能与冷冻过程中纺锤体和染色体的损伤有关,也可能与冷冻保护剂的残留毒性有关。此外,法律伦理问题也是卵母细胞冷冻保存技术面临的一大挑战。不同国家和地区对卵母细胞冷冻保存的法律和伦理规定各不相同,这在一定程度上限制了该技术的普及和应用。美国ICSI纺锤体Oosight Meta纺锤体在细胞分裂完成后迅速解体,为细胞质分裂提供空间。
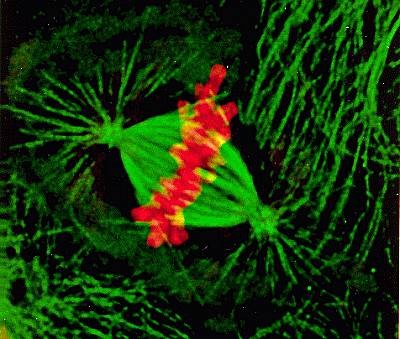
纺锤体在有丝分裂中发挥着至关重要的导航作用,其主要功能包括:排列与分裂染色体:纺锤体的完整性决定了染色体分裂的正确性。在细胞分裂中期,染色体在纺锤丝的牵引下,自动在赤道板排列整齐。当细胞进入分裂后期,纺锤体微管收缩,将染色体牵引至两极,形成两组数目相等的姐妹染色单体。这一过程确保了遗传信息的准确传递,避免了染色体分离错误导致的遗传异常。决定胞质分裂的分裂面:在染色体分裂的同时,纺锤体中的一部分微管不随染色体分裂到两极,而是停弛在纺锤体中间形成纺锤中心体。纺锤中心体的中心区域为两组极性相反的微管交叠区,称为纺锤中心区,它决定了接下来的胞质分裂面。胞质分裂开始于分裂后期的较晚期,一般结束于分裂末期后1-2小时,此期间两个子细胞由中心颗粒体连接。纺锤体通过精确控制胞质分裂面的位置,确保了细胞分裂的对称性和稳定性。
染色体当细胞从间期进入有丝分裂期,间期细胞微管网络解聚为游离的αβ-微管蛋白二聚体,再重组成纺锤体,介导染色体的运动;分裂末期纺锤体微管解聚,又重组形成细胞质微管网络。可分为:动粒微管:连接染色体动粒于两极的微管。极间微管:从两极发出,在纺锤体中部赤道区相互交错的微管。星体微管:中心体周围呈辐射分布的微管。染色体的运动依赖纺锤体微管的组装和去组装。在这一过程中动粒微管与动粒之间的滑动主要是依靠结合在动粒部位的驱动蛋白和动力蛋白沿微管的运动来完成。极微管在纺锤体中部交错,有些分布在极微管之间特殊的双极马达蛋白,其中2个马达蛋白沿一条微管运动,另2个马达结构域沿另一条微管运动。由于2条微管分别来自二极,故极性相反。当双极驱动蛋白四聚体沿微管向正极运动时,纺锤体二极间距离延长。反之纺锤体距离缩短。纺锤体在细胞分裂中的稳定性对于细胞存活至关重要。
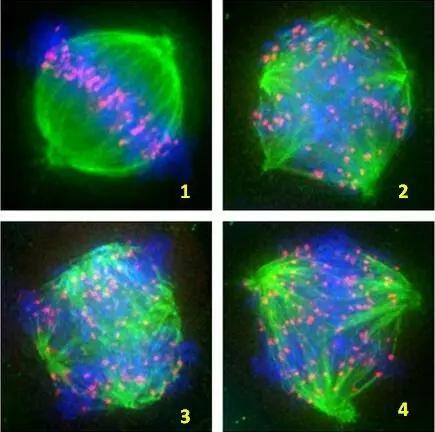
阿尔茨海默病患者中,微管蛋白(如tau蛋白)的突变和异常磷酸化会影响微管的稳定性和纺锤体的组装,导致染色体分离异常和细胞周期紊乱。纺锤体功能障碍会导致染色体不稳定,增加基因组的不稳定性,进而影响神经元的正常功能和存活。正常情况下,成熟的神经元处于G0期,不会重新进入细胞周期。然而,阿尔茨海默病患者中,神经元可能会重新进入细胞周期,但由于纺锤体功能障碍,无法完成正常的细胞分裂,导致细胞凋亡。在神经元中,纺锤体的正常功能对于神经元的发育、分化和维持至关重要。纺锤体微管的排列和稳定性受到细胞骨架的支撑。昆明无损观察纺锤体液晶偏光补偿器
纺锤体在细胞分裂完成后迅速解体,为细胞进入下一个周期做准备。美国克隆纺锤体卵质量评估
微管蛋白的突变会影响微管的聚合和解聚,导致纺锤体结构异常。例如,某些疾病中,微管蛋白的突变会导致纺锤体功能障碍,增加染色体非整倍性的风险。动粒与微管结合能力下降:动粒是染色体与纺锤体微管连接的关键结构,其功能障碍会影响染色体的正确捕捉和分离。例如,某些基因突变(如BUBR1突变)会影响动粒的功能,导致染色体分离错误。动粒通过信号传导途径与纺锤体检查点相互作用,确保染色体的正确分离。动粒信号传导异常会导致纺锤体检查点失效,增加染色体非整倍性的风险。美国克隆纺锤体卵质量评估